The Biological Evolution of Pain
B.Contestabile First version 2007 Last version 2024
Table of Contents
1. Introduction
2. Basics
2.1 Pain
2.2 Evolution
2.3 The Emergence of Consciousness
2.4 The Emergence of Pain
3.1 Pain as an Adaptive Trait
3.2 Non-Adaptive Pain
3.3 Painless Evolution
3.4 Pain as a Mystery
4.1 Intensity
4.2 Quantity
4.3 Distribution
5.1 Randomness
5.2 Constraints
6. Conclusion
Starting point
The capability to feel pain increases (discontinuously) with evolution and seems not to be limited. The capability to feel pleasure increases as well, but pain cannot be compensated by pleasure across individuals.
Type of problem
What is the role of pain in evolution?
Why does it increase?
How does the distribution of pain evolve?
The role of pain in evolution
The fact that pain states are associated with damaging experiences is the result of natural selection. Pain can be an adaptive trait.
In other cases pain is only a by-product of natural selection. The same allele can produce senseless pain or superior adaptation depending on the combination with other alleles, i.e. it produces either handicapped or advantaged individuals with regard to biological fitness. If the result is a net reproductive advantage, then the allele will tend to increase in frequency in the population.
The increase of pain
Once pain improves the survival value, it starts to have an impact on evolution.
The increasing capability to feel pain has to do with the increasing importance of learning mechanisms. The importance of learning mechanisms increases with the lifetime of the creatures and with the complexity of the environment. The behavior of long-lived creatures is shaped by painful experiences acting on these learning mechanisms. A wide range of emotions enhances the capability to respond to the environment. A wide range and differentiation of emotions implies a high degree (intensity and duration) of pain. Intensity and duration measure the importance of the event and induce a corresponding long-term storage in the memory. Under these premises the capability to feel a high degree of pain is superior with regard to biological fitness
The distribution of pain
Certain kinds of pain are prevented or mitigated by the phenomenon of biological altruism.
The general trend is an increasing inequality (injustice) in the distribution of pain.
Cultural level
The question, if the biological trend continuous on the cultural level is investigated in
The Cultural Evolution of Suffering
Starting point
The capability to feel pain increases (discontinuously) with evolution and seems not to be limited. The capability to feel pleasure increases as well, but pain cannot be compensated by pleasure across individuals.
Type of problem
- What is the role of pain in evolution?
- Why does it increase?
- How does the distribution of pain evolve?
2.1 Pain
Definition
- Pain is an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage (Pain, Wikipedia). Pain is always a personal experience.
The relationship between the sensory and emotional experience is complex:
- A good book can lessen a toothache, and a hot probe hurts more when you feel sad.
- The pain killer Paracetamol not only reduces physical pain, but also the pain of social rejection [Thomson].
- In the broken-heart-syndrome similar areas of the brain are activated, as in the case of pain, and persons who are affected feel an acute chest pain [Stefan, 111].
- Although pain is defined as a sensory and emotional experience, it is traditionally researched and clinically treated separately from emotion [Gilam]. Concerning the classification of pain within emotions see (Emotion, Wikipedia).
Types of pain
1. Nociceptive pain. Nociceptors are the nerves that detect potentially noxious stimuli, and nociception is what they do for a living: they send reports about tissue state. Reports about tissue state represent the sensory experience of pain. The emotional experience of pain is brain-generated and based on many factors, including but not limited to nociception [Ingraham 2019]. If the factor nociception dominates, then one speaks of nociceptive pain.
2. Neuropathic pain arises from damage to the nervous system itself, either from disease or injury.
3. Nociplastic pain is a neurological dysfunction
a. where the brain’s pain center responds disproportionally to minor injuries or inflammations or
b. where it sends pain signals, even if there is no mechanical, chemical or thermal irritation, and no damage done to the nervous system [Lawton]. Example: Fibromyalgia.
Assessment
The major method used in human pain studies is (still) self-reporting.
- An elementary form rates pain on a scale from 0 to 10.
- A more sophisticated form is the McGill Pain Questionnaire, where the patients are confronted with a list of 78 words in 20 sections that are related to pain.
- A complementary option is qualitative sensing testing where you apply stimuli and ask the individual to indicate when they start to feel it. This allows you to compare an individual’s experience of pain against the average [Demming].
How reliable is self-reporting? People can lie or deceive themselves in relation to pain [Broom, 18].
For that reason, there is a tendency to assess objective data:
- Certain pain-related metrics can be recorded with a smartwatch and then combined with self-reports [Demming].
- Some researchers claim that pain intensity can now be measured objectively, using a technique for analyzing MRI scans [Hamzelou].
Links
2.1.2 Pain in Animals
Definition
Currently there is no consensus on the definition of pain in animals.
This discordance has contributed to the contentious debate about which species feel pain [Walters].
Pain is a sensory and emotional experience. Whereas the sensory experience is somewhat understood, the emotional experience is still a mystery (see chapter 3.4). We can only try to define conditions that must be met for the emergence of pain. Following Thomas Metzinger’s commented conditions [Metzinger 2017, 246-249]:
(1) Conscious experience
Only beings with consciousness can experience pain. Whereas the sensory experience of pain can be processed in the brain without conscious awareness, the emotional experience of pain is typically accompanied by conscious awareness. Consciousness is often described as the minds subjective experience.
(2) Phenomenal self-model
The self-model theory of subjectivity postulates that a phenomenal self-model is required for subjective experience. The perception of a self is an ongoing process within the self-model. In other words: the self-model provides the framework through which we experience consciousness. The self-model includes among others:
- the sense of ownership, the nontranscendable subjective experience that it is myself who is experiencing something.
- the sense of agency, the feeling that we are the ones initiating our actions
- sensory and emotional experiences
Neuroscientific research indicates that brain structures associated with emotions, like the limbic system (which belongs to the cerebral cortex), are closely connected to areas involved in self-referential processing like the prefrontal cortex. This suggests a neurobiological link between emotions and the construction of a self-model.
(3) Negative valence
Pain is created by states representing a negative value being integrated into the phenomenal self-model (…). This does not mean that the system itself must have a full understanding of what pain really means (in terms of cognitive, conceptual or linguistic competences): it suffices that it does not want to undergo this present conscious experience, that it wants it to end. Of course, to create the aforementioned phenomenal urgency of change, the mere representation of an expected negative utility may suffice. Prerequisite for the creation of positive and negative valences is a brain that interprets sensory experiences in terms of their survival value.
(4) Phenomenal transparency
Transparent phenomenal states make their representational content appear as irrevocably real, as something the existence of which you cannot doubt. What is inaccessible to conscious experience is the simple fact, that this experience is taking place in a medium. Therefore, transparency of phenomenal content leads to the subjective impression of immediacy.
Sentience
Above definition implies that only sentient organisms can experience pain.
Sentience is the capacity to feel, perceive, or experience subjectively. Eighteenth-century philosophers used the concept to distinguish the ability to think (reason) from the ability to feel (sentience).
- In modern Western philosophy, sentience is the ability to experience sensations (known in philosophy of mind as qualia). Sensations are sensory experiences combined with subjective experiences (like pain). The exact relationship between these two kinds of experiences, however, is still a subject of research.
- In Eastern philosophy, sentience is a metaphysical quality of all things that require respect and care.
(Sentience, Wikipedia)
Since we do not have access to the experiences of animals, pain must be assessed by behavior, anatomy, and physiology.
Assessment by behavior
Why is pain so intense? A false step will leave an organism writhing in pain, crying out for days, maybe weeks before healing sets in. The cry of an animal in distress quickly attracts predators (…). It would seem that natural selection would have favored less sensitive nerves or brain sensations (genesispark.com).
Knowledge of the selective pressures affecting the species is needed before behavioral responses to pain can properly be interpreted. The vocalization of pain is a trade-off with regard to natural selection:
- Animals which live socially, can help one other when attacked by a predator. Hence the loud vocalization of pain is adaptive.
- In species which can very seldom collaborate in defense the situation is different. The predators select weak individuals and vocalizations of pain might well attract predators.
If the vocalization offers a net reproductive advantage (as in humans, primates, dogs and pigs) it will tend to increase in frequency in the population, otherwise not (as in antelopes and sheep) [Broom, 19].
It is possible to have nociception without pain. People with complete spinal transection still exhibit nociceptive reflexes below the level of the injury. Therefore, exhibiting short-term or long-term behavioral responses to harmful stimuli doesn’t give sufficient evidence that an animal feels pain [Adamo].
Assessment by anatomy and physiology
- It is important to differentiate between nociception and pain because the latter always encompasses an emotional component and the IASP stresses that nociceptor activation is itself not pain [Smith].
- Peripheral anatomical and most physiological aspects of the pain system vary little amongst species. However, different vertebrate groups vary considerably in the locations of function. It is not logical to assume that, because an area which has a certain function in man is small or absent in another group of vertebrates, the function itself is missing [Broom, 19]
- A perception of pain can exist without the involvement of pain receptors, but so can visual or auditory perceptions exist without their receptors being involved. The pain system should be considered as a whole [Broom, 20].
- It is possible to have conscious sensory experiences (including nociception) without a sense of ownership. In pain asymbolia – a special case of a depersonalization disorder – nociceptive signals of bodily damage are not attributed to the self [Gerrans]. Persons with this disorder have the conscious sensory experience of pain (and can describe it) but lack the corresponding emotional experience (they do not flinch, grimace, or express distress). In humans pain asymbolia usually results from injury to the brain, but in species without a phenomenal self-model the lacking emotional experience could be a normal condition.
Assessment by a holistic approach
The pain system as a whole can be characterized as follows:
- Nociceptors
- Brain structures
- Pathways to higher brain structures
- Opioid receptors and substances
- The observation that analgesics reduce nociceptive response
- Avoidance learning
- Suspension of normal behavior
[Bateson]
Theses on fish
- Fishes cannot experience pain, fear, or other emotions [Rose 2002].
- Fish are consciously aware of pain [Braithwaite 2010].
- Recent research increasingly challenges the view that fishes cannot perceive pain [Segner 2012]
- Fish lack the necessary neurocytoarchitecture, microcircuitry, and structural connectivity for the neural processing required for feeling pain [Key 2016].
- Mammals, birds and reptiles all show signs of emotional responses, while fish and amphibians do not. The brains of higher vertebrates are also much richer in receptors for dopamine, the neurotransmitter most closely associated with reward pathways [Holmes 2017, 29].
- Studies in fish have shown that the biology of the nociceptive system is strikingly similar to that found in mammals. It is highly likely that fish experience pain [Sneddon 2019].
Theses on invertebrates
- Most invertebrates do not feel pain [Adamo 2019] [Senate of Canada 2003].
- The cephalopod class within the molluscs, particularly the coleoidea subclass (cuttlefish, squid, octopus) are probably capable of feeling pain. The same is true for crustaceans like prawns and crabs [Braithwaite 2010] [Stelling 2014].
- There are insects that feel something akin to what humans class as pain [Gibbons].
- It is virtually certain that some invertebrates (amoebas) are not sentient [DeGrazia 2002]
Summary
We have come a long way since Descartes, who argued that all non-human beings were merely automata, without self-awareness and incapable of feeling [Stelling 2014, 39].
Many researchers point to a line between vertebrates and invertebrates:
- As a general rule one can say that pain in vertebrates is certain (with some exceptions like fishes where the issue is disputed).
- As a general rule one can say that pain in invertebrates is uncertain.
o There are exceptions (like cephalopods and some insects) where pain is probable.
o In some species (like amoebas) it is virtually certain that pain does not exist.
Links
Some of the most discussed issues are the following:
o Pain in Cephalopods (a molluscan class)
o Pain in Crustaceans (an arthropod taxon)
Definition
- Evolution is change in the heritable traits of biological populations over successive generations. Evolutionary processes give rise to diversity at every level of biological organisation, including the levels of species, individual organisms, and molecules.
- The four most widely recognized evolutionary processes are natural selection, genetic drift, mutation and gene migration.
- Repeated formation of new species (speciation), change within species (anagenesis), and loss of species (extinction) throughout the evolutionary history of life on Earth are demonstrated by shared sets of morphological and biochemical traits, including shared DNA sequences. More than 99 percent of all species that ever lived on Earth are estimated to be extinct. Estimates of Earth's current species range from 10 to 14 million. (Evolution, Wikipedia)
History
In the mid-19th century, Charles Darwin formulated the scientific theory of evolution by natural selection, published in his book On the Origin of Species (1859). Evolution by natural selection is a process demonstrated by the observation that more offspring are produced than can possibly survive, along with three facts about populations:
- traits vary among individuals with respect to morphology, physiology, and behaviour (phenotypic variation)
- different traits confer different rates of survival and reproduction (differential fitness)
- traits can be passed from generation to generation (heritability of fitness)
Thus, in successive generations members of a population are replaced by progeny of parents better adapted to survive and reproduce in the biophysical environment in which natural selection takes place. This teleonomy is the quality whereby the process of natural selection creates and preserves traits that are seemingly fitted for the functional roles they perform. (Evolution, Wikipedia)
In the 1920s and 1930s the so-called modern synthesis connected natural selection, mutation theory, and Mendelian inheritance into a unified theory that applied generally to any branch of biology (…). The publication of the structure of DNA by James Watson and Francis Crick in 1953 demonstrated a physical mechanism for inheritance. Molecular biology improved our understanding of the relationship between genotype and phenotype. Advancements were also made in phylogenetic systematics, mapping the transition of traits into a comparative and testable framework through the publication and use of evolutionary trees. (Evolution, Wikipedia)
In light of discoveries made in recent decades, some biologists have argued for an extended evolutionary synthesis, which would account for the effects of non-genetic inheritance modes, such as epigenetics, parental effects, ecological and cultural inheritance, and evolvability (Evolution, Wikipedia).
The extended evolutionary synthesis considers amongst others the theory of punctuated equilibrium and the multilevel selection advocated by Stephen Jay Gould in the 1980s.
Current finding suggest that the focus on genetic mutations only captures part of the story of adaptive evolution – the slowly changing part [Laland, 42]
In this paper we concentrate on natural selection, because the emergence of pain is linked to adaptation by natural selection.
Natural selection
Under selection, individuals with advantages or "adaptive" traits tend to be more successful than their peers reproductively—meaning they contribute more offspring to the succeeding generation than others do. When these traits have a genetic basis, selection can increase the prevalence of those traits, because offspring will inherit those traits from their parents. When selection is intense and persistent, adaptive traits become universal to the population or species, which may then be said to have evolved.
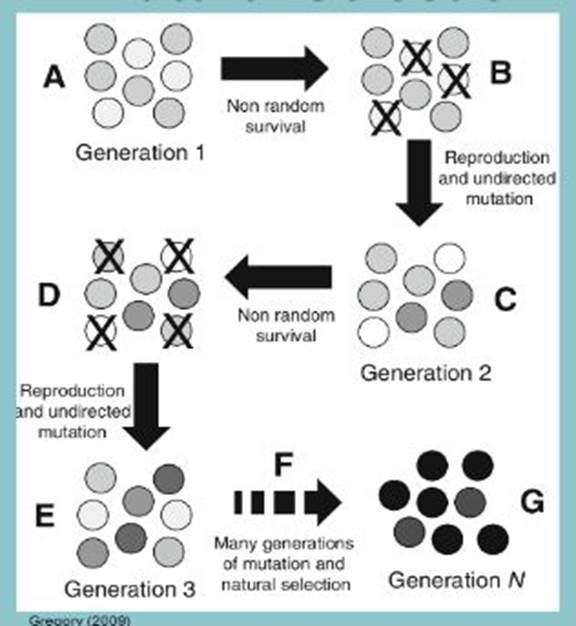
Diagram from the internet (author unknown)
Whether or not selection takes place depends on the conditions in which the individuals of a species find themselves. Adults, juveniles, embryos, and even eggs and sperm may undergo selection. Factors fostering selection include limits on resources (nourishment, habitat space, mates) and the existence of threats (predators, disease, adverse weather). Biologists often refer to such factors as selective pressures.
Subcategories of natural selection include sexual selection and ecological selection.
Selection occurs only when the individuals of a population are diverse in their characteristics—or more specifically when the traits of individuals differ with respect to how well they equip them to survive or exploit a particular pressure. In the absence of individual variation, or when variations are selectively neutral, selection does not occur.
Selection may act "negatively" as well as "positively."
- Negative selection or purifying selection decreases the prevalence of traits that diminish individuals' capacity to succeed reproductively (i.e. their fitness)
- positive selection increases the prevalence of adaptive traits.
Selection does not guarantee that advantageous traits or alleles will become prevalent within a population. Through genetic drift, such traits may become less common or disappear. In the face of selection even a so-called deleterious allele may become universal to the members of a species. This is a risk primarily in the case of "weak" selection (e.g. an infectious disease with only a low mortality rate) or small populations.
(Selection, Wikipedia).
Adaptation
Evolution influences every aspect of the form and behaviour of organisms. Most prominent are the specific behavioural and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding predators or attracting mates. Organisms can also respond to selection by cooperating with each other, usually by aiding their relatives or engaging in mutually beneficial symbiosis. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed (Evolution, Wikipedia)
- Adaptation is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats. Adaptation takes place in the life-cycle, which is often quite complex.
- Adaptation is one of the two main processes that explain the diverse species we see in biology (…) The other is speciation (species-splitting or cladogenesis), caused by geographical isolation or some other mechanism.
(Adaptation, Wikipedia)
An adaptive trait is the product (the bodily part) of the adaptation, i.e. those features of a species which result from the process of adaptation.
If and only if a trait fulfills the following criteria will evolutionary biologists in general declare the trait an adaptation:
1. The trait is a variation of an earlier form.
2. The trait is heritable through the transmission of genes.
3. The trait enhances (the probability of) reproductive success.
(Adaptationism, Wikipedia)
Some traits do not appear to be adaptive, that is, they appear to have a neutral or even deleterious effect on fitness in the current environment. Because genes have pleiotropic effects, not all traits may be functional (i.e. spandrels). Alternatively, a trait may have been adaptive at some point in an organism's evolutionary history, but a change in habitats caused what used to be an adaptation to become unnecessary or even a hindrance (maladaptations). Such adaptations are termed vestigial (Adaptation, Wikipedia)
2.3 The Emergence of Consciousness
Hierarchies
Antonio Damasio distinguishes between three levels of consciousness [Damasio]:
1) On the lowest level is the meta-representation of the body (the protoself), which is already present in primitive animals. This also includes the perception of the state of alertness. There is a continuous transition from alertness to inattentiveness, to sleep, to coma, and finally to death.
2) The next higher level is the so-called core-consciousness, the feeling for what happens, respectively the feeling that the proto-self is moving through the world and interacts with the world. Damasio describes the case of a patient who, for a period of 6 years, lived only with the core consciousness, i.e. in a world with no past and no future. The core-consciousness is also present in intelligent animals.
3) On the highest level is the extended consciousness, the link of the core-consciousness with autobiographical data, with a life story and its projection into the future. The extended consciousness exists only in humans and possibly in certain primates.
a) In contrast to the long-standing thesis that language is at the center of the extended consciousness, it seems to be the feelings, respectively the knowledge about having feelings. In strokes, patients may lose their speech. In the case of a cure, such patients say that their consciousness was completely intact, but that they simply could not communicate their condition.
b) An important role is also played by the connection of the brain’s left and the right hemisphere. If this connection is destroyed, then two consciousness-instances arise: one describes the world verbally, the other spatially. There is no communication between these two instances, but they are nevertheless represented by the same "ego-feeling".
A hierarchical structure with four levels of consciousness is proposed by [Kolodny, 3].
In vertebrates the levels of consciousness rely on two major brain systems [Fabbro]:
1. A “basal” subcortical system that includes the brainstem and the hypothalamus. This system may underpin the primary consciousness, which is likely present in all vertebrates. The subcortical system plays a key role in emotional processing and contributes to the sense of body ownership [Crucianelli].
2. A forebrain system that includes the medial and lateral structures of the cerebral hemispheres. This system may sustain the most sophisticated forms of consciousness.
The existence of emotions and a “sense of ownership” in invertebrates is a topic of ongoing research.
Dimensions
The idea that consciousness is structured in levels (with humans on top) is challenged by Jonathan Birch et.al [Birch]. He considers five separate dimensions of conscious experience:
1. Perceptual richness, the capability of an animal to distinguish details with each of its senses
2. Evaluative richness, the capacity to differentiate between positive and negative stimuli (analogous to the hedonic system of humans)
3. Integration: The extent to which an animal integrates the information from its sensory organs into a single experience.
4. Temporality: The capability to use information from the past and to plan for the future.
5. Selfhood: The ability to recognize one’s own mind and to understand that another animal has its own mind.
According to Birch and his colleagues, it doesn’t make sense to ask whether one animal is more or less conscious than another, since each species may score high on some of these facets of consciousness, but low on others [Robson 2021, 38].
In which stage of evolution did consciousness arise?
Thesis 1: Consciousness is a property of matter.
The mental cannot be reduced to the physical and somehow the property that, under special circumstances, enables a particular configuration of matter (e.g., the brain) to exhibit "consciousness" must be present in all matter, starting from the most fundamental constituents (Piero Scaruffi). The idea that the spiritual exists at the most elementary level can also be found in panpsychism, in particular in the philosophy of Albert North Whitehead.
Thesis 2: Consciousness emerged with life.
This thesis is advocated by the Cellular Basis of Consciousness theory:
The most primitive unicellular species of bacteria are conscious, though it is a sentience of a primitive kind. They have minds, though they are tiny and limited in scope. Bacteria, for example, have sophisticated sensory and perceptual systems, learn, form memories, make decisions based on information about their environment relative to internal metabolic states, communicate with each other, and even show a primitive form of altruism. All such functions are indicators of sentience [Reber].
The oldest evidence of life dates to around 3.5 billion years ago.
Thesis 3: Consciousness consists of bits of awareness
Visual qualia are quanta of visual awareness produced by circuits in the brain containing hundreds to thousands of neurons. Circuits with specific topologies reproducibly create the same type of qualia, and it is likely there are just a few diverse types. For example, there might only be three color qualia that produce the sensations of red, blue and green. When large numbers of these color qualia are combined in different ways, we create every possible color we humans can experience. Quantized visual awareness is consistent with what we know of biological systems. Basic sets of building blocks are used to create a huge variety of intricate structures in biology, and this occurs at every level of complexity. Imagine a single quale forming long ago within the nervous system of some simple organism. If this provided a selective advantage, over time the number and type of qualia would increase and eventually lead to complex forms of visual consciousness like that found in humans [Escobar].
The development of eyespots or rudimentary eyes likely occurred in some early multicellular organisms, such as flatworms, around 590 to 540 million years ago.
Let’s assume – in opposition to above theses – that consciousness requires a high level of complexity. The question then is when and how this level first emerged. Can we think of a single common ancestor? Or did it evolve independently in different lines, in much the same way that wings appeared separately in insects, birds and bats? An indication for the latter thesis is the finding, that in birds, the brain processing underpinning visual consciousness is located at a different part of the brain than in primates and humans [Robson 2021, 39].
Thesis 4: Consciousness can be explained by selective attention.
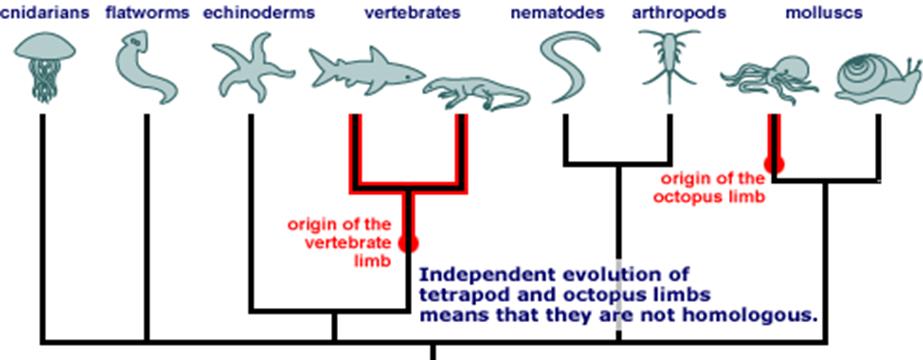
An animal is conscious, if it experiences the world subjectively, i.e. if it can make the distinctions “me, here, now”. Like hedonic valuation, subjective experience allows behavioral flexibility that goes beyond mere reflex responses. But it does not necessarily involve any of our more sophisticated abilities like emotion, reason or imagination. Subjectivity can be measured by selective attention, i.e. the ability to focus on just a few elements among all the sensory information available. Selective attention indicates that an individual is taking control of its perception and corresponds to an individual experience. The small brain of a fruit fly, for example, has a capacity for attention which could be the dawn of consciousness. Selective attention can be found in vertebrates, insects and octopuses at the very least. The common ancestor of these three groups was a very simple organism that resembled a flatworm (a simple bilaterian, unsegmented, invertebrate). Consciousness evolved separately in the three groups. All three groups feature nimble, fast-moving animals that encountered rapidly changing conditions as they moved. That puts a premium on flexible decision-making [Holmes 2017, 30-31].
With reference to the picture below: Insects belong to the arthropods, octopuses belong to the molluscs. Concerning the first appearance of these animals see [Marhall].
Picture from the internet (author unknown)
Insects were long thought to be simple, reflexive creatures with hardwired instinctual behaviors. No more [Koch]. Within the insects bees, for example, are not only remarkably smart but also kind of emotional. The inner lives of bees are governed by complex feelings – mental states that are in some ways similar to human emotions like discouragement and contentment [Schiffman, 41].
Thesis 5: Consciousness can be explained by unlimited associative learning [Jablonka].
It requires not only selective attention, but also the ability to knit multiple cues into a single perception that is more than the sum of its parts, and then use that compound cue to drive behavior. Furthermore, it requires the ability to distinguish between self and environment. Unlimited associative learning can be found in almost all vertebrates (except possibly lampreys), some arthropods such as insects and crustaceans, a few molluscs including octopuses and, perhaps, some snails and worms. If this thesis holds then consciousness evolved in early vertebrates and early arthropods during the Cambrian explosion, about 540 to 530 million years ago, when these groups diversified rapidly. Consciousness in octopuses probably evolved about 250 million years later, after their lineage diverged from other, less intellectually gifted mollusks such as clams and snails. The Cambrian explosion saw the emergence of most of the major animal groups alive today, and consciousness – driven by selection for powerful learning ability – might have helped drive that rapid evolution [Holmes 2017, 31].
Thesis 6: Consciousness can be explained by a mental model of attention.
To act on sensory information the animal requires a mental model of attention, for much the same reasons as it needs a mental model of its body (see Attention Schema Theory). This model is responsible for our conscious awareness of the world, and it may only be found in vertebrates [Holmes 2017, 31]. However, there is a mental model of the environment in spiders [Robson 2020], and spiders belong to the arthropods. On a side note: Some spiders can pick up sounds in the air using their webs as acoustic antennae [Wong, 15].
Conclusion
Consciousness is not clear-cut. There is no single dividing line between those species that enjoy the glow of an inner universe and those that don’t. There is not just one single way of being conscious. The animal kingdom is suffused with other kinds of minds and other kinds of consciousness, and they are not just mini versions of human consciousness [Holmes 2017, 31].
Origin of the nervous system
The evolution of the nervous system traces back to the earliest forms of life, with gradual complexity emerging over time. Sponges are the simplest multicellular animals and likely emerged more than 700 million years ago. They do not have a true nervous system, but they do have some sensory cells that can detect light and touch. These cells communicate with each other through gap junctions, which are channels that allow ions to pass from one cell to another. This mechanism helps coordinating the activities of different parts of the body. Gap junctions in sponges may be the precursors to synapses.
Diploblasts such as comb jellies are the next simplest multicellular animals and likely emerged more than 600 million years ago. Diploblasts have a simple nerve net that is spread throughout the body of the animal. This nerve net uses both gap junctions and synapses for communication between neurons. Gap junctions are good for quick and coordinated communication, while synapses are good for generating more complex patterns of signaling. Diploblasts can coordinate their movements, capture prey, and avoid predators. Cnidarians such as jellyfish, corals and sea anemones are the most complex diploblasts.
Origin of the brain
Prerequisite for the creation of a hedonic system is a brain that interprets sensory experiences in terms of their survival value. In the Precambrian world, around 600 million years ago, some animals evolved bilateral symmetry for the first time: that is, they now had a defined top and bottom, as well as a front and back. The current predominant view is that an ancestral bilaterian possessed a brain and centralized nerve cord [Strausfeld]. About 590 million years ago the bilateria underwent a profound evolutionary split. They divided into the protostomes and deuterostomes [Marshall].
- The deuterostomes eventually include all the vertebrates, plus echinoderms (starfish, sea urchins, sea cucumbers, etc.)
- The protostomes become all the arthropods (insects, spiders, crabs, shrimp, etc.), molluscs (snails, clams, octopuses, etc.), and various types of worms (flatworms, nematodes, earthworms, leeches, etc.)
The main part of this process occurred during the Cambrian explosion, about 540 to 530 million years ago. Utilizing comparative observations, the French naturalist Saint-Hilaire (1772-1844) was the first to propose that the ventral nervous system of arthropods corresponds to the dorsal nervous system of vertebrates.
Precursors of the hedonic system
Following two possible precursors of the hedonic system:
(1) There is a correspondence of action selection centers in arthropods and vertebrates, situated in the most anterior part of the brain. The selection of actions by these centers probably relied on circuits providing feed forward and feedback loops, modulated by dopamine signaling [Strausfeld]. Dopamine is known as a neurotransmitter to mediate reward and motivation, but it can also causally drive animals to avoid harmful situations and stimuli [Wenzel].
(2) Another possible precursor is nociception. All organisms have some form of nociception, even Bacteria. Leeches, which belong to the same phylum as flatworms, contain touch, pressure, and nociceptive cells [Vizioli]. There are three types of stimuli that nociceptors are sensitive to: thermal, mechanical, and chemical. A growing body of evidence suggests that thermal sensitivities predate the protostome–deuterostome split [Himmel].
There is a connection between the two mechanisms. Dopamine is an important modulator in nociception and analgesia.
Consciousness and emotion
Animals can survive solely based on sensory experiences. But emotional experiences (including pain) improve the biological fitness. A wide and flexible range of emotions enhances the capability to respond to the environment. It is plausible to assume that emotions emerged gradually, similar to the gradual emergence of consciousness. Concerning the relation between consciousness and emotions there are at least three remarkable theses:
- Thesis 1: Emotional and non-emotional (sensory) conscious experiences are not fundamentally different. What differs are the kinds of inputs that are processed by one central network of cognition in the cerebral cortex [LeDoux]. If thesis 1 holds, then there could be species with conscious sensory experiences and no emotions, like pain asymbolia in humans. The emergence of non-emotional consciousness could have preceded emotional consciousness. It may also be possible to construct intelligent robots with a similar kind of consciousness.
- Thesis 2: The upper brainstem plays a crucial role in our consciousness. This part of the brain helps us make decisions and motivates us to act. Mammals can still show goal-directed behavior even after their cerebral cortex has been removed experimentally. Children born without a cerebral cortex can still be conscious [Merker]. This challenges the traditional view, represented by thesis 1, according to which consciousness is solely a function of the cerebral cortex. The brainstem processes sensory information and triggers automatic emotional responses, such as the fight-or-flight response, without engaging the higher-order cognitive processes that are involved in more complex emotional experiences. This allows to respond to the environment in a timely manner, without having to think about it. In evolutionary terms the cerebral cortex is about 320 million years old, the brainstem about 520 million years.
- Thesis 3: Consciousness and emotions are inextricably linked:
There is no phenomenal awareness without feelings (feelings are how we interpret emotions). Feelings are an integral part of any conscious experience. And because basic phenomenal awareness necessarily includes feelings, so do all other types of consciousness that are built on top of it [Kolodny, 4].
This contradicts thesis 1, which says that there is a non-emotional kind of consciousness.
For information on the evolution of emotions see [Plutchik].
Pain and the sense of touch
It is crucial for understanding the emergence of pain that it cannot be replaced by the sense of touch. One might expect the sense of touch to warn an animal of thorns, because tactile cells are highly sensitive sensors with excellent spatial resolution, quite adapted to thorn-shaped objects. However, the sense of touch is not suitable as a sense of alarm, for two main reasons:
1. The sensory quality of touch is the exploration of objects, the analytical study of shape, motion, and temperature.
2. The sense of touch requires attention, the conscious turning to the object under investigation.
Both properties are unsuitable for an alarm sense. Exploration takes too long, and an alarm sense cannot presuppose attention; its purpose is rather to generate attention. The pain sense is therefore independent of all other senses in structure and function; it has its own sensory cells (nociception), its own regulatory mechanisms, and its own access to reflexes, motivations, and consciousness [Frings].
In which stage of evolution did pain arise?
For a neurological view on the evolution of pain see [Broom, 20].
For a description of pain mechanisms across species see [Walters].
Thesis 1: Pain emerged in early vertebrates
The conditions for pain, as described in chapter 2.1.2 were first fulfilled in early vertebrates, i.e. in the Cambrian explosion, ca.540 to 530 million years ago. All the different vertebrate species are endowed with a common brain archetype [Fabbro].
Thesis 2: Pain emerged in the first fully terrestrial vertebrates
The first fully terrestrial vertebrates, the common ancestors of modern reptiles, birds, and mammals, emerged ca. 300 million years ago. These ancestors faced challenges that its aquatic cousins did not, like temperature regulation and water conservation. In this new environment hedonistic valuation was a big advantage, because it improved the adaptability of the organism [Holmes 2017, 29-30].
The terrestrial environment, however, is not a sufficient explanation, because pain also emerged independently within crustaceans and molluscs [Crook] and probably also in some insects. It is even possible that pain in molluscs preceded pain in vertebrates.
Are there better worlds than the human world?
- Candidates for a better world are species with a hedonic system but less intense experiences. Let us assume the hedonic scale in animals is non-linear and asymmetric, as it is with humans [Caviola, 5-6]. In this case the balance of pleasure and pain could be better in the animal kingdom, because the most intense pleasure that is eliminated counts less than the most intense pain that is eliminated. Disquieting, however, is the fact that the hedonistic balance of animals, even if better than the human one, could still be negative. Reasons for this concern can be found in chapter 4.2.
- A different idea is based on the hypothesis that non-emotional consciousness emerged earlier in evolution than emotional consciousness. If that is true, then there is an immense world of consciousness in a hedonistically neutral state. Many of the early life forms (or similar forms) still exist.
In both cases one could argue that the richness of human lives cannot be replaced by a better hedonistic balance. Following two counterarguments:
1. Animals do not miss the richness of human lives. They cannot suffer from something they don’t know.
2. The richness of positive human experiences is devalued by the richness of negative experiences.
Let’s assume that the animal kingdom is preferable, indeed. Under these premises there are excellent chances to exist in a better world. Take the world of insects as an example. There are ca. 1018 insects and less than 1010 humans, so that the chance to exist as a human is minimal. For compassionate people, however, this is no reason to be reassured. Pain cannot be compensated by pleasure across individuals, no matter how many happy individuals exist. The phenomenon of extreme pain and suffering remains the ultimate challenge of suffering-focused ethics.
Is the emergence of pain a necessary event in evolution? For an answer to this question see chapter 5.
Once the phenomenon of pain arose, it proved to be valuable in the struggle for survival. A function, which does not provide an evolutionary advantage, disappears again with time, because it consumes energy unnecessarily.
Definition
Pain is an adaptive trait if it enhances the probability of that organism to reproduce.
Example:
The pain of a prey being wounded by a predator is adaptive, because it makes the prey flee. But there is no evolutionary pressure to stop the pain of a dying prey:
“It is easy to imagine a. gene that, say, tranquillizes gazelles when they are about to suffer a killing bite. Would such a gene be favored by natural selection? Not unless the act of tranquillizing a gazelle improved that gene's chances of being propagated into future generations. It is hard to see why this should be so and we may therefore guess that gazelles suffer horrible pain and fear when they are pursued to the death - as most of them eventually are (River out of Eden, Richard Dawkins).
The proliferation of pain
The pain system has evolved and is probably old in evolutionary terms. Most aspects of the pain system, including the feeling of pain, are functional and adaptive. There is evidence for the pain system in all vertebrates which have been studied and there is evidence for some aspects of it in invertebrates. The similarities in the pain system across vertebrates are considerably greater than the differences, except in respect of behavioral responses to pain which vary adaptively according to the way of life [Broom, 10].
The selective advantage of pain
Subjective sensations, such as pleasures and pains, can be adaptive traits of organisms.
So far as we know, there is no fundamental physical reason why pleasure is attached to some circumstances and pains to others. We might have evolved to find the sensation of having a finger caught in a door intensely pleasurable, just as dung-flies may well relish the smell of excrement. On the evolutionary hypothesis, the fact that pleasures are generally associated with beneficial, and pains with detrimental, experiences, is the result of natural selection among random variations: those individuals who happen to have an association of this kind have higher biological fitness than those who have no such association, or the reverse association (No Pain - no Gain by David B.)
Emotions are set to maximize Darwinian fitness [Nesse 1991, 37].
Concerning the asymmetry between pain and pleasure see Antinatalism and the Minimization of Suffering.
If individuals with “the capability to feel pain” have a higher biological fitness then – conversely – those without this capability have a selective disadvantage. Example: People with congenital insensitivity to pain [Nagasako] have a reduced life expectancy.
The rare individuals who cannot feel pain fail even to experience discomfort from staying in the same position for long periods. Their unnatural stillness impairs the blood supply to their joints, which then deteriorate. Such pain-free people usually die by early adulthood from tissue damage and infections [Nesse 1998].
The range of phenomena
Human beings do experience pain, and there is a correlation between incidents of pain and tissue damage.
- Either this trait is present in the human phenotype because it was selected for, or
- its presence is a byproduct of selection for some other trait.
The first thesis is more plausible: Pain accompanies too wide a range of physical phenomena to be a byproduct of selection for something else [Saidel]
Example: A squid may feel pain very differently from a human. Shortly after a squid’s fin is crushed, nociceptors become active not only in the region of wound, but across a large part of the body. This suggests that if it feels pain then rather than being able to pinpoint the location of the wound, an injured squid may hurt all over. This makes sense insofar, as a squid (in contrast to an octopus) is not able to tend the wound. An all-over sensitivity with regard to touch and visual stimuli keeps the squid more alert and wary [Stelling, 41].
Pain is analogous to a “smoke alarm”
An obstacle to perceiving the benefits of defences arises from the observation that many individuals regularly experience seemingly worthless reactions of anxiety, pain, fever, diarrhoea or nausea. The explanation requires an analysis of the regulation of defensive responses in terms of signal-detection theory. A circulating toxin may come from something in the stomach. An organism can expel it by vomiting, but only at a price. The cost of a false alarm— vomiting when no toxin is truly present— is only a few calories. But the penalty for a single missed authentic alarm— failure to vomit when confronted with a toxin—may be death. Natural selection therefore tends to shape regulation mechanisms with hair triggers, following what we call the smoke-detector principle. A smoke alarm that will reliably wake a sleeping family in the event of any fire will necessarily give a false alarm every time the toast burns. The price of the human body’s numerous “smoke alarms” is much suffering that is completely normal but in most instances unnecessary [Nesse 1998].
Pain makes learning more effective
In honest signaling theory a cost is being paid by the system to facilitate a reliable selection of courses of action. It is important to realize that signaling can be carried out not only between but also within individuals. The concepts of signaling and honesty or reliability are thus directly applicable to the questions of the evolutionary origins and role of pain [Kolodny, 6].
By analogy to the distinction between retributive and consequentialist approaches to justice, it would be useless for pain to be merely a punishment for a poor choice made in the past: it must somehow affect future behavior. Moreover, pain cannot be merely another informational aspect of the process of credit assignment for the purposes of learning. Pain is functionally orthogonal to such outcome dependent rewards: it is the cost of bidding on participation in action, whose evolutionary role stems from the need to ensure the reliability of the bids. Specifically, in vertebrates this functionality is implemented by a distributed network of “actors” that receive performance feedback from a “critic” mechanism. To the extent that actors compete against each other for access to behavioral control, there is a need for a mechanism that would promote honest interactions within the action selection and learning system. Affective experiences reflect the transactions that promote honesty — and thereby make learning more effective — by imposing up-front cost on the actors. The price paid in the currency of pain thus helps elevate those actors that can afford it and that are therefore more likely to propel the system that embodies them “through the hardships to the stars” — per aspera ad astra [Kolodny, 11].
Example: Consider a situation in which you have spied some blackberries deep in a bramble thicket and are attempting to reach for them. The present pain caused by the thorns and the memories of past pain inflicted by thumbtacks and nails motivate you to withdraw; yet, something inside you tells you to persevere, and so you push through until the sweet objective is attained. It is now time to assess your behavior. The assessment is not just of the reward (how sweet the blackberries turned out to be) but of its relationship to your original confidence and to whatever damage you incurred in acting on it. Next time, your confidence in deciding to persevere in the face of pain will be higher, or lower, depending on the outcome of the assessment. The outcome of this assessment is distinct from the reward prediction error in reinforcement learning, because confidence in the sense proposed here has to do not just with the expected reward but also with what it takes to achieve it [Kolodny, 6-7].
The unique pain of being human
Pain researcher Patrick Wall was the first person to suggest a functional view of pain – that it should be understood as a mixture of sensation and the motivation to make it stop, not sensation alone. His starting point was the now well researched placebo effect (…). Several stimuli and situations that are not painful in animals have come to be experienced as painful for humans. This is because our obvious distress elicits help from others and hence offers a survival advantage [Finlay, 28].
Example: Labor pain appears only in humans. Primates appear to give birth with little pain. Ungulates such as horses and cattle produce large, long-limbed offspring with a substantial chance of complications, but with little evidence of distress. Any such evidence, in fact, could prove fatal by alerting predators [Finlay, 29].
Recent theses
Migraine and cluster headache seem to make no evolutionary sense. Recent research, however, suggests that
- migraine is linked to sickness behavior, which is an adaptive response to internal and external stressors, characterized by withdrawal and lethargy
- cluster headache is linked to the fight-or-flight reaction, which is an adaptive response to external threat, characterized by motor restlessness and agitation.
[Bonavita]
Supernormal stimuli
The resemblance between the intensity of pain and the importance of the risk is not perfect, because natural selection can be a poor designer. The result is often glitches in animals, including supernormal stimuli. Such glitches help explain pains which are not, or at least no longer directly adaptive. (Pain, Wikipedia)
Also the pain resulting from a kidney stone stuck in the ureter is non-adaptive [Bateson].
Chronic pain
The severity and chronicity of some pain has always been hard to understand in the context of evolution, particularly when it actually interferes with function. Our first experience with being burned tells us to never, ever touch fire again, and we don’t and (this is the important part) we’re definitely better off for it. But when the message of pain is actually debilitating for long periods, how exactly is that sensation helping? Were cavemen with chronic back pain better hunters or something? (…) It turns out that we have a system that purposefully takes pain to the next level. It didn’t have to be that way. It was just an accident of biology, another design flaw.
The system to blame here is the infection-fighting system, the immune system, blindingly fast, agile, fierce cells bristling with biological weapons (…). A few billion of these little monsters – called neutrophils – are your best friends every time there is a genuine risk of infection — which there definitely is every time the skin is broken (…) But neutrophils have no concept of a false alarm. Internal injury — sterile tissue damage, any injury where there is exactly zero risk of infection — causes exactly the same reaction. Neutrophils rush to the scene and start doing their thing. They attack and kill any cells in the area — ours included — just in case (…). The problem is that they have a bogus alarm signal.
In a well-designed biology, the neutrophils would know the difference between injury with a risk of infection and an injury without a risk of infection. Viewed strictly as an engineering problem, this is a no-brainer: there are countless candidate chemical signatures that could do the trick, “smells” that are exclusive to each kind of damage. But the neutrophils are oblivious to those smells, because there is only one signal that does matter to them, just one telltale sign that crops up inside our very own cells, as well as on actual invaders. That’s because there’s actually an “invader” inside every single cell in our body: a welcome guest providing a critical service, without which we would literally die in seconds. However, that guest is still foreign, technically. It still smells like an invader — like bacteria, specifically.
A long, long time ago, multicellular organisms started a marriage of convenience with bacteria. Those bacteria became permanent residents of every cell in our bodies. They became, in fact, cellular organs, vital subsystems (…). They produce energy, and a great deal of it, like a power plant. Mitochondria have always stayed true to themselves, remarkably autonomous symbionts. They even still retain their own DNA. Our immune systems evolved relatively separately from the mitochondria, which remained safely tucked away inside our cells. As long as the mitochondria stay inside our cells, there’s no trouble. But when cells are damaged and mitochondria spill out into our tissue fluids, the neutrophils attack, because mitochondria look like invaders to neutrophils (…).
But neutrophils don’t even mind killing our own cells. In fact, it’s their job. They are indiscriminate better-safe-than-sorry killers that are only too happy to take out a few you-cells in the heat of battle. “It’s the only way to be sure.” And so not only is this component of inflammation completely pointless in the absence of any actual infection hazard, but healthy cells are actually killed. (…) It means that pain and injury actually get worse for no reason whatsoever [Ingraham 2015]
Idiopathic pain
Idiopathic pain (pain that persists after the trauma or pathology has healed, or that arises without any apparent cause), may be an exception to the idea that pain is helpful to survival, although some psychodynamic psychologists argue that such pain is psychogenic, enlisted as a protective distraction to keep dangerous emotions unconscious (Pain, Wikipedia)
Trade-offs
Compromise is inherent in every adaptation. Arm bones three times their current thickness would almost never break, but Homo sapiens would be lumbering creatures on a never-ending quest for calcium. More sensitive ears might sometimes be useful, but we would be distracted by the noise of air molecules banging into our eardrums.
Such trade-offs also exist at the genetic level. If a mutation offers a net reproductive advantage, it will tend to increase in frequency in a population even if it causes vulnerability to disease. People with two copies of the sickle cell gene, for example, suffer terrible pain and die young. People with two copies of the “normal” gene are at high risk of death from malaria. But individuals with one of each are protected from both malaria and sickle cell disease. Where malaria is prevalent, such people are fitter, in the Darwinian sense, than members of either other group. So even though the sickle cell gene causes disease, it is selected for where malaria persists. Which is the “healthy” allele in this environment? The question has no answer. There is no one normal human genome—there are only genes [Nesse 1998].
Painless creatures
A flexible brain can improve the biological fitness but it does not necessarily lead to superiority over other creatures:
- Three-quarters of all known animals are insects, more than a billion for each person on the planet. More than half of the insect families survived the most catastrophic mass extinction the world has ever seen [O’Donoghue].
- Although complex species have evolved, they occur as a side effect of the overall number of organisms. The overwhelming majority of species are microscopic prokaryotes, which form about half the world's biomass despite their small size, and constitute the vast majority of Earth's biodiversity. Simple organisms have been the dominant form of life on Earth throughout its history and continue to be the main form of life up to the present day, with complex life only appearing more diverse because it is more noticeable (Evolution, Wikipedia)
Prokaryotes can be divided into Archaea and Bacteria. Bacteria are very efficient regarding survival despite lacking intelligence. They dispose of a CRISPR mechanism which destroys invading viruses [Li]. Furthermore, due to their flexible genome, they can permanently create new forms and adapt to the environment. There is a competition between the survival strategy of flexible genomes and the strategy of flexible brains, and the outcome of this race is open:
o Bacteria always find ways to outwit the defensive strategies of the human body and the human medicine
o Medical research is permanently expanding its knowledge to figure out the bacteria’s tricks.
[Schatz]
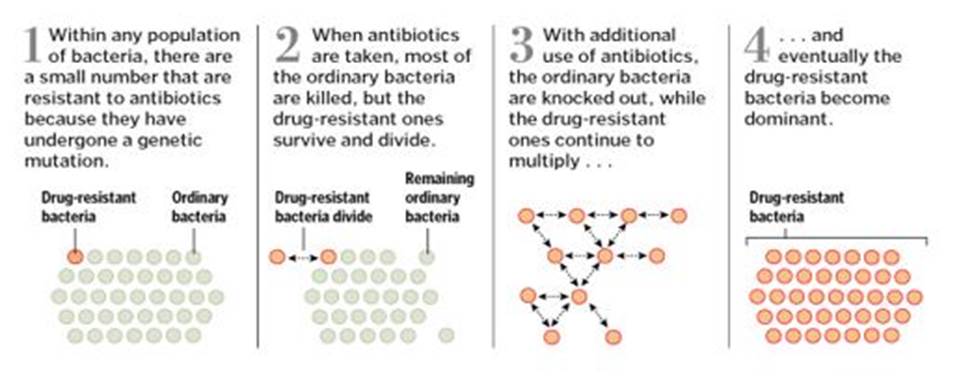
Graphic from the internet (Author: Washington Post)
The selective advantage of painlessness
First of all painless evolution is the original and most probable type of evolution. If sentient life emerges, however, then there may be an evolutionary advantage in preserving painlessness with regard to certain parts or functions of the body:
In cases where pain does not improve or even worsens the biological fitness, pain should not emerge (or re-disappear) in the course of evolution.
Examples confirming this thesis:
- The desert mole rat has a highly unusual set of physical traits that enables it to thrive in an otherwise harsh, underground environment, including a lack of pain sensation in its skin.
- Grasshopper mice (Onychomys torridus) are resistant to the painful, and potentially lethal, stings of one of their menu items – the Arizona bark scorpion (Centruroides sculpturatus).
- The human brain has none of the three types of pain receptors. Neither the mechanical, thermal nor chemical sensors would be triggering before the underlying cause would already cause major damage to the extremely sensitive structures of the brain. Thus omitting them in favor of a tighter brain structure not disrupted by other neurons is an advantage (Wesley Crusher)
Why is pain phenomenally felt and not merely informational?
- The standard account of pain, which holds it to be a signal sent by a damage-monitoring process to the central controller, fails to explain why pain is hurtful and not just informational [Kolodny, 5]. Consider the withdrawal reflex. Disengagement from a noxious stimulus is enacted immediately and does not, in principle, require that the stimulus also be experienced as painful. Thus, the role of pain here cannot be merely to motivate an immediate withdrawal, which is in fact taken care of by the reflex arc. Neither can the role of pain be merely to instill later aversion to the stimulus: aversion, like motivation, is informational and does not have to involve phenomenal experience [Kolodny, 6].
- Could pain play a role in reinforcement learning? By definition, learning from behavioral outcomes requires that the blame or credit for the results of an action be apportioned to its causal components, giving rise to the so-called Credit Assignment Problem that is central to AI (and to the birth of New Connectionism). Clearly, credit assignment is an informational function, and so, just like motivation and aversion, it does not need phenomenal pain to mediate it [Kolodny, 6].
- Could pain play a role in the context of honest signaling theory? The answer is yes (see chapter 3.1). Affective experiences reflect the transactions that promote honesty (and thereby make learning more effective) by imposing up-front cost on the actors. The price paid in the currency of pain thus helps elevate those actors that can afford it. But why must the currency be pain? Can’t the system pay the price while remaining emotionally indifferent? According to the Dynamical Emergence Theory all phenomenality, including pain and other affective processes, is equated with certain intrinsic topological properties of the system’s trajectories through its state space. Whatever the dynamics of pain turns out to be, it must be intrinsic: it is not enough for the system’s states to be interpreted externally as meeting the stipulated conditions [Kolodny, 9].
The evolutionary function of consciousness is still unclear. Given the close relation between affects and consciousness (chapter 2.3) it is plausible to assume that the mystery of pain will not be revealed before the hard problem of consciousness is solved.
Learning mechanisms
The emergence of higher degrees of pain is not an accidental by-product of evolution; it is the consequence of the adaptive function of pain.
The increasing capability to feel pain has to do with the increasing importance of learning mechanisms.
The importance of learning mechanisms increases
- with the lifetime of the creatures and
- with the complexity of the environment.
Lifetime
Pain is realised in biology as an educational tool:
- Vertebrates are relatively long-lived creatures and learning shapes their behaviour.
- Invertebrates are short-lived and their behaviour is largely genetically determined. There is less pressure for the evolution of pain here.
(No brain, no pain? The Hindu, Mar 10, 2005)
There is also a different pressure for the evolution of pain within the invertebrates, depending on the lifetime of the creatures.
Example: Crustaceans like prawns and crabs show responses to injuries that go beyond reflex – prolonged and complicated behavior, which clearly involves the central nervous system. They are capable of learning, just as you would expect to see from an animal that experienced pain. They are even able to weigh different needs when responding to a noxious stimulus [Stelling, 40].
Crustaceans have a similar-sized nervous system like fruit flies, but unlike crustaceans, insects seem to have no pain-related behaviours. Why is that?
Neurons consume a lot of energy and there is evolutionary pressure to keep brains as compact as possible. There need to be good reasons to develop a brain that is capable of feeling pain [Stelling, 41]
The reasons to develop such a brain are lacking in the world of insects:
If an animal’s lifespan is not long enough to benefit from learning – as is the case with most insects – then pain has no use [Stelling, 41].
Complexity of the environment
In his book, The Greatest Show on Earth: The Evidence for Evolution, biologist Richard Dawkins grapples with the question of why pain has to be so very painful. He describes the alternative as a simple, mental rising of a "red flag". To argue why that red flag might be insufficient, Dawkins explains that drives must compete with each other within living beings. The most fit creature would be the one whose pains are well balanced. Those pains which mean certain death when ignored will become the most powerfully felt. The relative intensities of pain, then, may resemble the relative importance of that risk to our ancestors (lack of food, too much cold, or serious injuries are felt as agony, whereas minor damage is felt as mere discomfort). (Pain, Wikipedia)
The behaviour of long-lived creatures is shaped by painful experiences acting on learning mechanisms. A wide range and differentiation of emotions enhances the capability to respond to a complex environment. A wide range of emotions includes a high degree (intensity and duration) of pain. Intensity and duration measure the importance of the event and induce a corresponding long-term storage in the memory. Under these premises the capability to feel a high degree of pain is superior with regard to biological fitness
Cultural level
The question, if the biological trend continues on the cultural level is investigated in
The Cultural Evolution of Suffering
There is a biological pressure to expand populations at the cost of the quality of life:
1.
Populations grow faster than
resources:
The total amount of suffering per year in
the natural world is beyond all decent contemplation. During the minute that it
takes me to compose this sentence, thousands of animals are being eaten alive,
others are running for their lives, whimpering with fear, others are being
slowly devoured from within by rasping parasites, thousands of all kinds are
dying of starvation, thirst and disease. It must be so.
If there is ever a time of plenty, this very fact will automatically lead to an increase in population until the natural state of starvation and misery is restored (River out of Eden, Richard Dawkins).
2. Nature cares about species, not about individuals. Example:
Sea turtles usually lay around 100 eggs at a time, but on average only one of the eggs from the nest will survive to adulthood (…) hatchlings may be eaten within minutes of hatching as they make their initial run for the ocean (Sea Turtle, Wikipedia).
The expansion of sentient life is tied to an expansion of pain. Pleasure expands as well, but pain cannot be compensated by pleasure across individuals.
There are two opposing trends in the distribution of pain:
1. Evolution is a process of increasing differentiation, implying increasing inequalities. Not only are the various forms of life are unequal but also the individual members within each type. Individual inequalities increase with the evolution of the species. Complex organisms differ more from each other than simple organisms. Evolution creates unequal distributions within all dimensions of life, in particular unequal distributions of pain. With the degree of pain, the degree of injustice increases as well:
In a universe of electrons and selfish genes, blind physical forces and genetic replication, some organisms are going to get hurt, others are going to get lucky, and you won't find any rhyme or reason in it, nor any justice. The universe that we observe has precisely the properties we should expect if there is, at bottom, no design, no purpose, no evil and no good, nothing but pitiless indifference [Dawkins 1995, 80-85]
2. Certain kinds of pain (e.g. the pain of starving or being attacked by predators) are prevented or mitigated by altruism:
Donor-recipient correlation, rather than genetic relatedness, is the key to the evolution of altruism. What is needed for altruism to evolve, is (…) that the recipients of altruism have a greater than random chance of being fellow altruists. Whether this higher than random donor-recipient correlation arises because partners tend to be relatives, or because altruists are able to seek out other altruists and choose them as partners, or for some other reason, makes no difference to the evolutionary dynamics (Biological Altruism, Stanford Encyclopedia of Philosophy).
The net result of these two opposing trends is an increasing injustice in the distribution of pain.
Is the emergence of pain a necessary event in evolution? Or would replaying evolution yield a different result?
A common misconception is that evolution has goals, long-term plans, or an innate tendency for "progress" as expressed in beliefs such as orthogenesis and evolutionism (Evolution, Wikipedia)
According to Stephen Jay Gould there is no inherent drive towards long-term progress. Uncritical commentaries often portray evolution as a ladder of progress, leading towards bigger, faster, and smarter organisms, the assumption being that evolution is somehow driving organisms to get more complex and ultimately more like humankind. Gould argued that evolution's drive was not towards complexity, but towards diversification. Because life is constrained to begin with a simple starting point (like bacteria), any diversity resulting from this start, by random walk, will have a skewed distribution and therefore be perceived to move in the direction of higher complexity. But life, Gould argued, can also easily adapt towards simplification, as is often the case with parasites. (Stephen Jay Gould, Wikipedia)
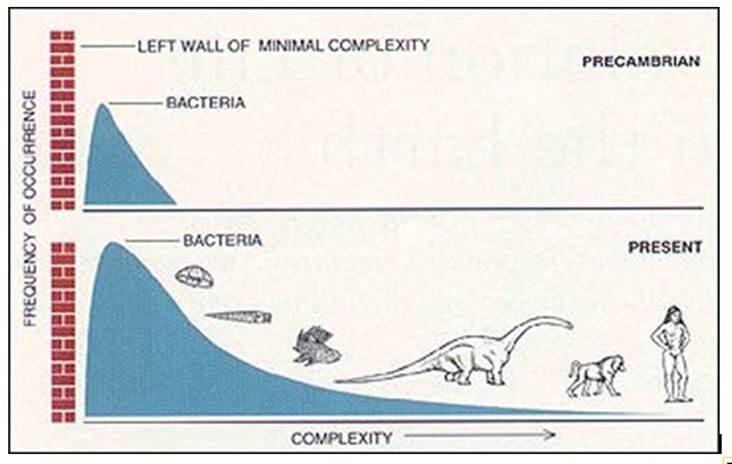
Picture from the internet (author unknown)
According to Gould, evolution would produce very different outcomes every time, if life could be started again. The study of evolution therefore resembles the study of history. It is something we can only understand in retrospect. The course of evolution is often shaped by accidental events:
- The fittest do not always survive. If it hadn’t been for an asteroid strike, for instance, we mammals might still be scurrying about in mortal fear of dinosaurs
- Despite many claims to the contrary, most of the human genome is just junk. This junk has accumulated because natural selection has not been strong enough to remove it.
- In small populations even mutations that are slightly harmful can spread throughout the population simply by chance (see Genetic drift).
- Speciation is almost always an accidental process.
[Holmes 2015]
Dawkins’ gene-centered view
Richard Dawkins emphasizes the role of adaptation in evolution. If the environment is reflected in the DNA, then the result of replaying evolution cannot be random.
The utility function of life, that which is being maximized in the natural world is DNA survival. But DNA is not floating free: it is locked up in living bodies and it has to make the most of the levers of power at its disposal. (…) Each change is due to evolution: “Each generation is a filter, a sieve: good genes tend to fall through the sieve into the next generation; bad genes tend to end up in bodies that die without reproducing. (…) Animals are survival machines – robot vehicles blindly programmed to preserve the selfish molecules known as genes” (Dawkins, 2007).
- God's utility function (also called biological utility function or utility function of life) is a phrase coined by Richard Dawkins in his book River Out of Eden (…). Dawkins uses this phrase to expound the Gene-centered view of evolution by equating the phrase to the meaning of life or the purpose of life. This is the why question about life which philosophers and theologians have been pondering in vain for ages, and is a counterpart to the how question about nature which engineers have been able to resolve successfully (…)
- Dawkins rephrases the word purpose in terms of what economists call a utility function, meaning "that which is maximized". Engineers often investigate the intended purpose (or utility function) of a piece of equipment using reverse engineering. Dawkins uses this technique to reverse-engineer the purpose in the mind of the Divine Engineer of Nature, or the Utility Function of God.
- According to Dawkins, it is a mistake to assume that an ecosystem or a species as a whole exists for a purpose. In fact, it is wrong to suppose that individual organisms lead a meaningful life either (…). The unit of selection is the gene, not an individual, or any other higher-order group as championed by proponents of group selection. In nature, only genes have a utility function – to perpetuate their own existence with indifference to great sufferings inflicted upon the organisms they build, exploit and discard (God’s Utility Function, Wikipedia).
Gould’s criticism
Gould views selection as working on many levels, and has called attention to a hierarchical perspective of selection. Gould also called the claims of Selfish Gene "strict adaptationism" and describing them as excessively "reductionist". He saw the theory as leading to a simplistic "algorithmic" theory of evolution, or even to the re-introduction of a teleological principle (…) To Gould, the fatal flaw was that "no matter how much power Dawkins wishes to assign to genes, there is one thing that he cannot give them – direct visibility to natural selection.” Rather, the unit of selection is the phenotype, not the genotype, because it is phenotypes that interact with the environment at the natural-selection interface. So, in Kim Sterelny's summation of Gould's view, gene differences do not cause evolutionary changes in populations, they register those changes.”
Richard Dawkins comments on what he sees as the main difference between his position and that of the late Stephen Jay Gould. He concludes that it is his own vision that genes play a causal role in evolution, while Gould sees genes as passive recorders of what worked better than what (Gene-centered view, Wikipedia)
Convergent evolution
Richard Dawkins insists that although mutations may be random, evolution is not. If we look at how evolution has turned out on neighboring islands, then we see the constraints to randomness. There are only limited ways of flying and swimming, for instance, which is why wings and fins have independently evolved on many occasions [Holmes 2015].
Example: On different Caribbean islands (Cuba, Jamaica, Hispaniola and Puerto Rico) the Anolis lizards have adapted to similar niches (ground, trunk, tree-crown, tree-top) and developed similar feet, extremities and tail lengths. The similar-looking lizards, however, are not closely related to each other. Closely related are the dissimilar-looking lizards, which live on the same island [Blount, 41]. For more examples see Convergent evolution.
Partial rerun
What would be the result of replaying evolution? In a limited, partial sense the tape of life has been rerun many times. Very different species living in similar environments have independently evolved in similar ways. Many outcomes of evolution are not accidental but inevitable [Chorost, 36].
Catastrophes only delay or accelerate the developmental process, but they do not significantly change it. If the dinosaurs weren’t killed by a meteorite they would have survived until Earth’s next glaciation. The cold would have killed off those dinosaurs living north and south of the tropics, opening up niches for the warm-blooded mammals and birds that coexisted with them. Eventually tool-users not unlike us would have evolved and sooner or later any dinosaurs remaining in the tropics would have been hunted to extinction. While the outcome of evolution is not fully determined, it is powerfully constrained. Direction and constraint, however, do not imply design and purpose [Chorost, 37].
Complete restart
What would be the result of replaying evolution from beginning?
- The emergence of complex cells depended on a highly unlikely merger of two kinds of simple cells. Bacteria-like life forms could therefore be expected, but not more sophisticated organisms.
- Assumed that life did get past this hurdle, one could expect to see photosynthesisers and predators, parasites and decomposers. Big-brained primates are highly unlikely but some other brainy, social animal could take over the planet. There’s clearly an adaptive zone in most habitats that involves intelligence. And it has become clear that many traits we once thought of a uniquely human, from language to tool-making, exist to some extent in many other animals [Holmes 2015].... in particular the capability to experience pain.
The role of pain in evolution
- The fact that pain states are associated with damaging experiences is the result of natural selection. Pain can be an adaptive trait.
- In other cases pain is only a by-product of natural selection. The same allele can produce senseless pain or superior adaptation depending on the combination with other alleles, i.e. it produces either handicapped or advantaged individuals with regard to biological fitness. If the result is a net reproductive advantage, then the allele will tend to increase in frequency in the population.
The increase of pain
- Once pain improves the survival value, it starts to have an impact on evolution.
- The increasing capability to feel pain has to do with the increasing importance of learning mechanisms. The importance of learning mechanisms increases with the lifetime of the creatures and with the complexity of the environment. The behaviour of long-lived creatures is shaped by painful experiences acting on these learning mechanisms. A wide range of emotions enhances the capability to respond to the environment. A wide range and differentiation of emotions implies a high degree (intensity and duration) of pain. Intensity and duration measure the importance of the event and induce a corresponding long-term storage in the memory. Under these premises the capability to feel a high degree of pain is superior with regard to biological fitness
The distribution of pain
- Certain kinds of pain are prevented or mitigated by the phenomenon of biological altruism.
- The general trend is an increasing inequality (injustice) in the distribution of pain.
Cultural level
The question, if the biological trend continuous on the cultural level is investigated in
The Cultural Evolution of Suffering
1. Adamo Shelley A. (2019), Is it pain if it does not hurt? On the unlikelihood of insect pain, Cambridge University Press
2. Bateson Patrick (1991), Assessment of pain in animals, Animal Behavior 42, 827-839.
3. Birch Jonathan (2020), Dimensions of Animal Consciousness, Texas A&M University, available from https://eeb.tamu.edu/files/2020/09/Birch-et-al-2020.pdf
4. Blount Zachary D. (2018), Evolution zwischen Zufall und Wiederholung, Spektrum der Wissenschaft, March, 38-47
5. Bonavita V., De Simone R. (2011), Pain as an evolutionary necessity, Neurological Sciences, Vol.32, Supplement 1, 61-66
6. Braithwaite Victoria (2010), Do Fish Feel Pain?, Oxford University Press
7. Broom, D.M. (2001), The Evolution of Pain, University of Cambridge, available from www.vdt.ugent.be/sites/default/files/art70103.pdf
8. Caviola Lucius, David Althaus, Andreas Mogensen & Geoffrey Goodwin (2021), Population ethical intuitions, Department of Psychology, Harvard University
9. Chorost Michael (2012), The Ascent of Life, New Scientist, 21 January, 35-37
10. Crook Robyn J. (2021), Behavioral and neurophysiological evidence suggests affective pain experience in octopus, Science Direct
11. Crucianelli Laura et.al. (2023), Subcortical contributions to the sense of body ownership, Brain, 17.October, online ahead of print, Oxford Academic
12. Damasio Antonio (1999), The Feeling of What Happens: Body and Emotion in the Making of Consciousness. New York: Harcourt Brace.
13. Dawkins Richard (1995), God's Utility Function, Scientific American, November, 80-85
14. Dawkins Richard (2007), Study Notes
15. DeGrazia David, What Animals are Like
16. Demming Anna (2022), How can you measure someone’s pain, New Scientist, 9 November, 43
17. Douglas Kate (2010), How Powerful is the Subconscious?, New Scientist, 3 April
18. Escobar W.A. (2013), Quantized visual awareness, Frontiers in Psychology
19. Fabbro Franco, S.Aglioti, M.Bergamasco, A.Clarici, J.Panksepp (2015), Evolutionary aspects of self- and world consciousness in vertebrates, Frontiers in Human Neurosciences
20. Feinberg Todd E. and Jon Mallatt (2013), The evolutionary and genetic origins of consciousness in the Cambrian Period over 500 million years ago, Frontiers in Psychology
21. Finlay Barbara (2015) The unique pain of being human, New Scientist, 9 May, 28-29
22. Frings Stephan (2021), Der Schmerzsinn, in: Die Sinne der Tiere, 519-540, available from www.link.springer.com
23. Gerrans Philipe (2020), Pain Asymbolia as Depersonalization for Pain Experience, Frontiers in Psychology
24. Gibbons Matilda et.al. (2022), Can insects feel pain? A review of the neural and behavioral evidence, Advances in Insect Physiology, Volume 63, 155-229
25. Gilam Gadi, James J. Gross, Tor D. Wager, Francis J. Keefe, Sean C. Mackey (2020), What Is the Relationship between Pain and Emotion? Bridging Constructs and Communities, Neuroview, Vol.107, Issue 1, 17-21
26. Hamzelou Jessica (2010), The scanner that feels your pain, in New Scientist, 6 March, p.6
27. Himmel Nathaniel J., Jamin M. Letcher, Akira Sakurai, Thomas R. Gray, Maggie N. Benson and Daniel N. Cox (2019), Drosophila menthol sensitivity and the Precambrian origins of transient receptor potential-dependent chemosensation, The Royal Society Publishing
28. Holmes Bob (2015), Life Chances, New Scientist, 14 March, 32-35
29. Holmes Bob (2017), Why be conscious?, New Scientist, 13 May, 29-31
30. Ingraham Paul (2015), Why Does Pain Hurt?, PainScience.com
31. Ingraham Paul (2019), The 3 Basic Types of Pain, PainScience.com
32. Jablonka Eva, Simona Ginsburg (2019), The Evolution of the Sensitive Soul, MIT Press
33. Key Brian (2016), Why fish do not feel pain, Animal Sentience 3(1)
34. Koch Christoph (2008), Exploring Consciousness through the Study of Bees, Scientific American, 1 December
35. Kolodny Oren, Roy Moyal, Shimon Edelman (2021), A possible evolutionary function of pain and other affective dimensions of phenomenal conscious experience, in The Neuroscience of Consciousness, Special Issue: Consciousness Science and its Theories
36. Laland Kevin (2016), Evolution Evolves, New Scientist, 24 September, 42-45
37. Lawton Graham (2022), The Third Type of Pain, New Scientist, 9 November, 41
38. LeDoux Joseph and Richard Brown (2017), A higher-order theory of emotional consciousness, available from www.pnas.org
39. Li Gege (2021), Taming CRISPR, Evolutionary arms race, New Scientist, 16 January, 44
40. Marshall Michael (2009), Timeline: The evolution of life, New Scientist, 14 July
41. Merker Bjorn (2015), Consciousness without a cerebral cortex: A challenge for neuroscience and medicine, Cambridge University Press
42. Metzinger Thomas (2009), Der Ego Tunnel, Berlin Verlag
43. Metzinger Thomas (2017), Suffering, the cognitive scotoma, in The Return of Consciousness, pp.237-262, Editors Kurt Almqvist & Anders Haag, Stockholm, Sweden
44. Nagasako E.M., Oaklander A.L., Dworkin M.H. (2003), Congenital Insensitivity to Pain: an Update, Pain 101, 213-219, Elsevier
45. Nesse Randolph M. (1991), What Good is Feeling Bad? The Sciences, Nov./Dec.
46. Nesse Randolph M., Williams George C. (1998), Evolution and the Origins of Disease, Scientific American, November
47. O’Donoghue James (2015), Small Wonders, New Scientist, 7 November, 36-39
48. Plutchik Robert (1962), The Emotions, Revised Edition, University Press of America. For a visualization see Evolution of Emotions
49. Reber, A. S. (2019), The First Minds: Caterpillars, Karyotes, and Consciousness. New York: Oxford University Press.
50. Robson David (2020), Spiders think with their webs challenging our ideas of intelligence, New Scientist, 8 February, 44-45
51. Robson David (2021), What is consciousness like in other animals?, New Scientist, 10 July, 38-39
52. Rose James D. (2002), The Neurobehavioral Nature of Fishes and the Question of Awareness and Pain, CRC Press
53. Saidel Eric (2007), The Evolution of Pain, An Argument for the Causal Efficacy of Mental States, University of Southwestern Lousiana
54. Schatz Gottfried (2011), Planet der Mikroben, Warum wir Infektionskrankheiten nie endgültig besiegen werden, Neue Zürcher Zeitung Nr.274 vom 23.Nov, S.53
55. Schiffman Richard (2018), Buzz Off, New Scientist, 9 June, 40-41
56. Segner Helmut (2012). Fish.Nociception and Pain – A Biological Perspective. Ethics commission of the Swiss confederation.
57. Senate of Canada, Standing Committee on Legal and Constitutional Affairs (2003), Bill C-10B (Cruelty to Animals), June.12, Do Invertebrates Feel Pain?
58. Smith Ewan, Lewin Gary (2009), Nociceptors: a phylogenetic view, Journal of Comparative Physiology, 195(12): 1089–1106.
59. Sneddon Lynne U. (2004), Evolution of nociception and pain: evidence from fish models, The Royal Society Publishing
60. Stefan Cornelia (2017), Homo Patiens. A socio-philosophical concept, Ph.D.Thesis Nr.726023, Università degli studi dell ‘Insubria, Italy
61. Stelling Tamar (2014), Alien Pain, New Scientist, 22 February, 38-41
62. Strausfeld Nicholas J., Frank Hirth (2015), Introduction to ‘Origin and evolution of the nervous system’, The Royal Society Publishing
63. Thomson Helen (2022), What are the boundaries between emotions and pain?, New Scientist, 9 November, 40
64. Vizioli Jacopo, Francesco Drago, Christophe Lefebvre (2016), Neuroprotection and Immunity in the Medicinal Leech Hirudo medicinalis: What About Microglia?, Science Direct, Lessons in Immunity, 67-78
66. Wenzel Jennifer M. et.al. (2018), Phasic Dopamine Signals in the Nucleus Accumbens that Cause Active Avoidance Require Endocannabinoid Mobilization in the Midbrain. Current Biology, Cell Press, Cambridge, USA
67. Wong Carissa (2022), Spiderwebs may be the world's most sensitive ears, New Scientist, 19 February, 15